棘皮動物をモチーフとするポケモンの形態についての考察
ご挨拶
線虫亭線虫(以下、「著者」)と申します。この記事では、棘皮動物をモチーフとするポケモン達のデザインがモチーフの特徴をどのように反映しているのかを、棘皮動物に特徴的なボディプランの観点から見ていきます。なお、この記事におけるゲームのスクリーンショットは全て匿名の協力者より提供されたものです。この場を借りて御礼申し上げます。
以下常体。
- ご挨拶
- この記事の目的と方針
- 棘皮動物のボディプラン
- 考察の進め方
- ウミユリとウミユリモチーフのポケモン達
- ヒトデとヒトデモチーフのポケモン達
- ナマコとナマコモチーフのポケモン
- ウニとウニモチーフのポケモン
- 後書き
- 参考文献
この記事の目的と方針
この記事の目的の一つは、後述する「棘皮動物のボディプラン」について、それがどのように棘皮動物をモチーフとするポケモン(以下、「棘皮ポケモン」)のデザインに反映されているのか(いないのか)を考察することである。より砕けた表現をすれば、棘皮ポケモンの形態の棘皮動物「らしさ」について考察する。この記事では、あるポケモンが現実の生物の系統関係のどこに位置するかをポケモンの形態から推論することや、それによってポケモンどうしの近縁関係を議論することは行わない。棘皮ポケモンのモチーフはいずれも形態以外の要素、即ち名前や『分類』、図鑑説明文から推定したものであり、この推定と形態がどのように整合するかをこの記事では問題とする。また、ポケモンのデザインがモチーフとなる生物の形態的特徴を反映しているかどうかと、そのデザインが「良い」かどうかは別であり、この記事では後者の評価は控える。
この記事のもう一つの目的は、これからポケモンの形態を考察する人々が参照しやすいように整理した考察を、誰でも参照できる形で、文字媒体として保存することである。ポケモンの形態を少しでも考察したことがあるという人は多いだろうが、そのアイデアは頭の中にあるか親しい誰かに話すか、ツイート(古いものは一見すると消えてしまうので参照しにくい)となるかという場合が多いだろう。考察を記事として整理し参照可能な形で保存しようとすることは労力がかかるためか、人々が行った考察全てを文献として参照できる状況にはなっていない(昨今ではアイデアを出し合う様子を動画として配信して保存する場合もあるが、良し悪しは別にしてそのような場合考察が整理されているわけではない)。幸い今の著者には労力をかけるだけの資源があるため、一考察者として整理された考察を保存しようと思った次第である。この記事の中には「車輪の再発明」と呼ばれるアイデアも多く存在するだろうが、読者の皆様においては、この記事の目的を考慮してご容赦いただきたい。
棘皮動物のボディプラン
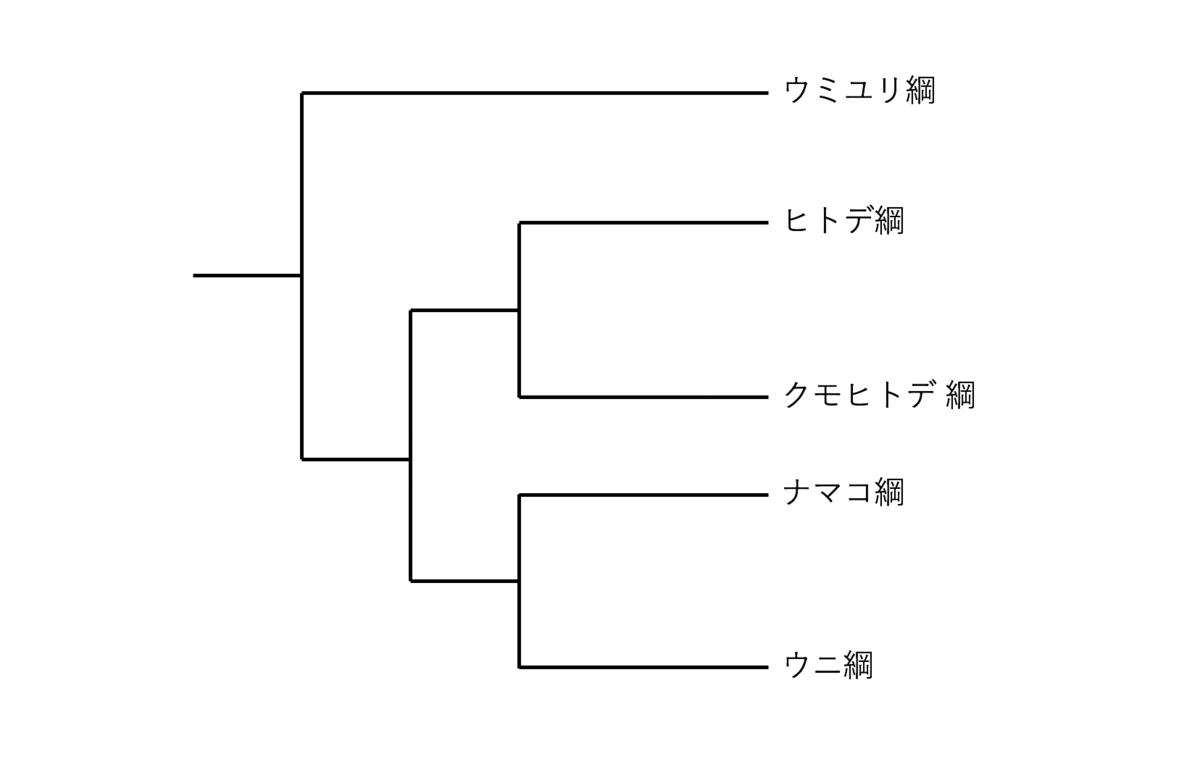
図1:現在考えられている棘皮動物の現生5綱の系統関係。図はReich et al.(2015)を参考に作成した。
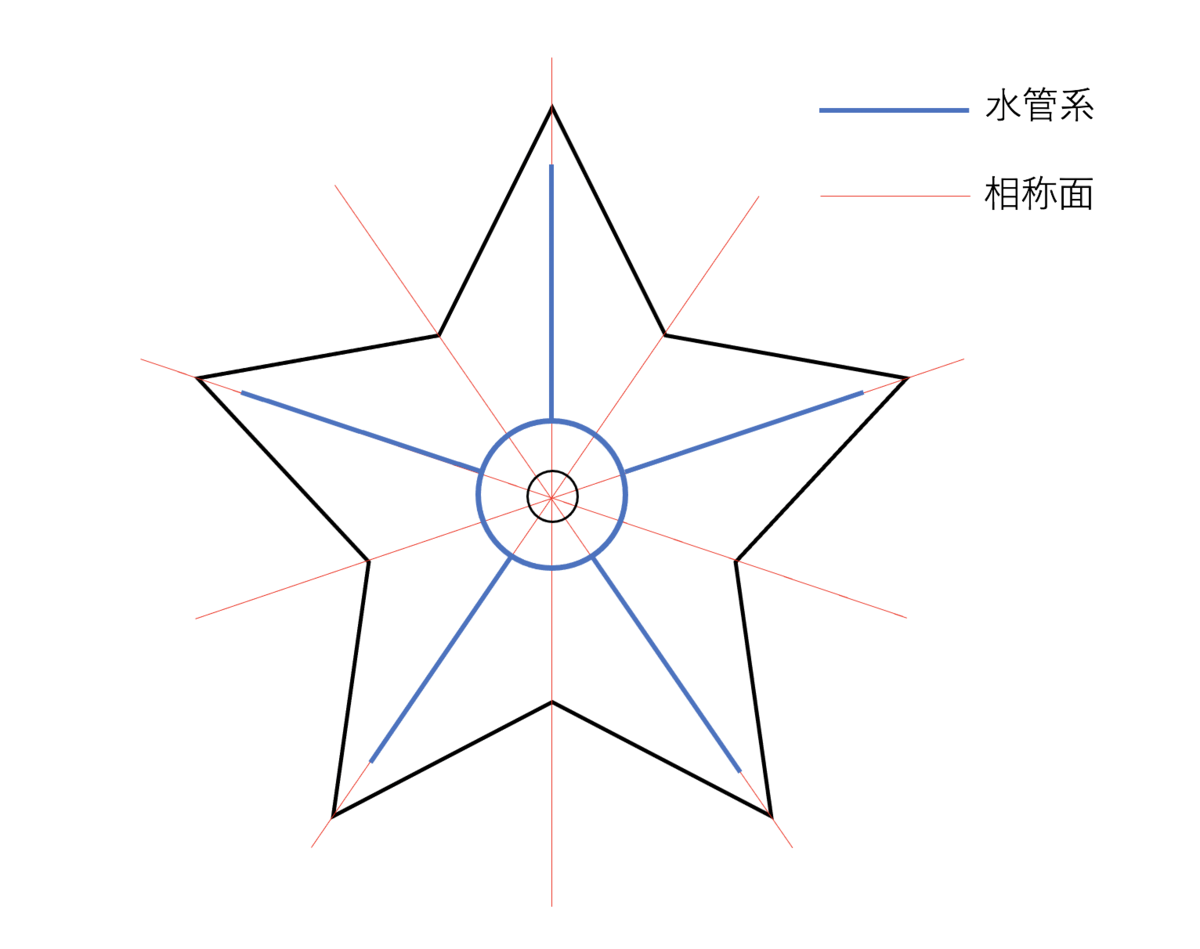
棘皮動物は後生動物(以下、「動物」)の一門である棘皮動物門に含まれる生物のことで、現生のものはウミユリ綱、ヒトデ綱、クモヒトデ綱、ナマコ綱、ウニ綱の5綱に分類される。現生棘皮動物は単系統群であり、綱どうしの系統関係を図1に示す。現生棘皮動物一般に共通する身体構造上の特徴として、「口側-反口側軸を中心軸とする(五)放射相称性」と「水管系の存在」が挙げられる。以下、それぞれについて詳しく述べる。
口側-反口側軸を中心軸とする(五)放射相称性について
棘皮動物、特にその成体の特徴としてよく挙げられることと言えば五放射相称(種や個体によっては五ではない場合もある)のボディプランだが、外部構造や内部構造の配置から考えるとその中心軸は口を通ると考えて差し支えないだろう(図2)。外形としては口を起点として放射状に歩帯(綱によって歩帯溝や食溝などと呼ばれる場合もある)が伸び、その周囲に間歩帯が存在している形態をとる。内部構造としては、口の周囲を環状水管が取り巻きそこから放射相称のそれぞれのユニット(大抵は5つ)に向かって放射水管が伸びる水管系や、それに並行して走る神経系がある。このような形態は棘皮動物門の現生5綱全てで共通しており、棘皮動物に特徴的なボディプランであると言える(ただしどの綱でも、幼生の形態は左右相称である)。なお放射相称の体を持つ棘皮動物は、ナマコと一部のウニを除き移動の際の前後軸が定まらない(360度どの向きにも移動できる)。棘皮動物の体軸を表す用語としては、「口側-反口側」軸が用いられる。口側が口のある側、反口側がその反対側である。
水管系について
口を中心とする放射相称の体を持つ動物としては他に刺胞動物門(イソギンチャクやサンゴを含む動物門)や有櫛動物門(クシクラゲやクラゲムシを含む動物門)があるが、棘皮動物にはこれらの動物とは異なり水管系が存在する。水管系は体腔由来の器官系で、内部に海水を取り込みそれを用いて呼吸や排出などを行う循環系の役割を持つと同時に、その一部が体外へと伸びて管足となり、身体の移動や餌の口への運搬に用いられる。水管系の大部分は体内に存在するため外部形態として観察できる部分は限られる。一つは先述の管足であり、これは放射水管の一部が体外へと露出しているものである。もう一つは多孔板と呼ばれる外部の海水と水管系を繋げる穴が多く開口した部分で、ヒトデやウニで見られる。
この記事では上記の2点について、即ち「口側-反口側軸を中心軸とする(五)放射相称性があるかどうか」及び「水管系に関する形態的特徴があるかどうか」を基準として棘皮ポケモンのデザインを考察する。
考察の進め方
棘皮動物をモチーフとするポケモンはヒトデマン・スターミー(ヒトデ綱)、リリーラ・ユレイドル(ウミユリ綱)、ナマコブシ(ナマコ綱)、ヒドイデ・ドヒドイデ(ヒトデ綱)、バチンウニ(ウニ綱)の計8種族である(()内はモチーフとなる綱)。これらのポケモンについて、モチーフとなる綱毎に分け、それぞれの綱について次のように進める。まず、対象とする綱の生物の形態的特徴を「口側-反口側軸を中心軸とする(五)放射相称性」及び「水管系に関する形態的特徴」の観点から概説する。次に、対象とする綱をモチーフとするポケモンについて、写真を用いながら口側-反口側軸を定める(なお、口の位置は「ソード・シールド」のキャンプでカレーを食べる映像や、「ポケモンGO」できのみを食べる映像から推定した)。最後に、対象とするポケモンのデザインがモチーフとなる綱の生物の形態的特徴をどのように反映しているかを、「口側-反口側軸を中心軸とする(五)放射相称性があるかどうか」及び「水管系に関する形態的特徴があるかどうか」の観点から述べる。なお、ポケモンの身体の部位の表記については図鑑の記述に従い、図鑑に表記されていない名称を用いる場合は「(**と考えられるもの)」などといった形でその旨を記した。また、文中に登場するポケモンの部位の名称は画像写真にその位置と共に記載した。
ウミユリとウミユリモチーフのポケモン達
ウミユリについて
ウミユリ綱には有茎のウミユリ類と無茎のウミシダ類が含まれるが、ここではウミユリ類について説明する。ウミユリの身体は冠部(萼と腕に分けられる)、萼の反口側から繋がる茎部、茎部先端にある根部に分けられる。萼には神経系をはじめとする諸内臓器官が収められており、上面(口盤、上蓋)のほぼ中心に口が開く。口からは5本の食溝(歩帯溝)が放射状に伸び、萼から放射状に生える腕の根本にそれぞれ繋がる。食溝は腕や腕の両側に生える羽枝の先端にまで繋がり、その内側にある管足で小型プランクトンを捕獲し、口まで運搬する役割を担う。腕は先端に行くまでに何度も分岐し、基本的に5の倍数本あるように見える(ただし分岐の多い種では各腕で必ずしも一様に分岐しないので5の倍数にならない場合がある)。茎部は茎板と呼ばれる構造が靭帯によって連なってできており、その断面は五角形(ウミユリ目)または円形(ホソウミユリ目)である。ウミユリ目の場合、一部の茎板(節板と呼ばれる)からは五本の巻枝が輪生する(ホソウミユリ目の巻枝はほとんど失われている)。巻枝は根部と共に身体を支持するために用いられる器官で、これによって外界の構造物にしがみつく。根部は固着のために用いられる部位で、固着する環境によって円盤状、掌状、細かく分岐するものなど様々な形態をとる。以上から、腕、食溝、巻枝はウミユリの口側-反口側軸を中心軸とした五放射相称性がよく現れる外部形態であると言える。また、水管系は上蓋に点在する水孔によって外部の海水と接続する。従って、外部から観察可能な水管系は食溝の管足と水孔となる。
リリーラとユレイドル


リリーラとその『進化』先のユレイドルのモチーフは、『分類』や名前からウミユリであると推測される。
まずは触手に注目する。リリーラ・ユレイドル共に壷状の頭(と考えられる部分)から伸びる触手は8本、放射状に生えている。リリーラやユレイドルの頭はウミユリの萼に、触手はウミユリの腕に対応すると考えられるが、ウミユリの場合は先述の通り萼から5の倍数本の腕が放射状に生えるのに対し、リリーラやユレイドルの触手の本数は5の倍数とはなっていない。ユレイドルの図鑑説明(ウルトラムーン)には「8つのしょくしゅで えものを とらえ しょうかえきで とかしながら くう。」とあり、単に触手が少なめに描画されているわけではない可能性が高い。従ってリリーラやユレイドルの触手の本数は、ウミユリの五放射相称性についての特徴を反映したデザインではないと言える。
次に首と体に注目する。リリーラやユレイドルの体からは5つの突起が生えており、歩く時にこれら(特に左右の4つ)を脚のように動かす。これらの突起と体はポケモンの身体全体を支持する役割を持っていると言える。ウミユリで身体全体を支持するのに用いられる部位は茎部と茎部から生える巻枝と根部であり、リリーラやユレイドルの首と体と突起はウミユリのこれらの部位に対応していると考えられる。先述のようにウミユリの巻枝は(生える場合は)口側-反口側軸を中心軸とした五放射相称に生えるが、リリーラやユレイドルの体の突起の生え方も(完全な相称ではないものの)口側-反口側軸を中心軸とした五放射相称に近い配置となっている。位置関係としてはリリーラやユレイドルの体はウミユリの根部との方が茎部とよりもよく対応するものの、反口側の外部形態にも口側-反口側軸を中心軸とした放射相称性が見られるという点で、リリーラやユレイドルの体のデザインはウミユリの五放射相称性をやや反映していると言える。
最後に頭(と思しき部分)に注目する。先述のようにリリーラやユレイドルの頭が対応するウミユリの萼には、その口側に口とそこから放射状に5本伸びる食溝が存在する。食溝には外部形態として観察できる水管系の一部である管足が並んでいる。しかしリリーラやユレイドルの頭の内側は黒く描写されており、より詳細に観察することができない。また、ウミユリの食溝は先述のように腕や羽枝の先にも繋がるが、ウミユリの腕に対応するであろうリリーラやユレイドルの触手には食溝らしき構造は観察できない。さらに、頭には水孔のような穴が点在する様子も見られない。従って、リリーラやユレイドルの頭と触手のデザインは水管系に関するウミユリの外部形態の特徴を反映しているとは言えない。
ヒトデとヒトデモチーフのポケモン達
ヒトデについて
ヒトデの身体は中心部の盤とそこから放射状に生える腕に一応分けられるものの、両者に厳密な区別はない。腕の数は種や個体によって異なり、5本の種も5本以上の種(オニヒトデでは11-18本)もある。口側の中心には口が開き、口を中心として腕の先端まで放射状に伸びる歩帯溝が存在する。歩帯溝には2列又は4列に管足が並び、ヒトデはこれを用いて移動や外界の構造物への付着、摂食を行う。盤に存在する環状水管は石管(石灰質の管)によって盤の反口側にある多孔板と連絡している。多孔板は小さな穴を多く持つ石灰質の小円盤で、この穴を通して外界と水管系の間で海水をやり取りする。多孔板の数は種によって異なり、腕の数とは比例しない。以上から、腕や歩帯溝の配置の仕方はヒトデの口側-反口側軸を中心軸とした放射相称性がよく現れる外部形態であると言える。また、管足やそれが並ぶ歩帯溝、及び多孔板は外部から観察可能な水管系の一部である。
ヒトデマンとスターミー

ヒトデマンとその『進化』先のスターミーのモチーフは、名前からヒトデであると推測される。
体の中心にあるコア(とヒトデマンの場合コアから伸びて反口側にまで達するリング状のもの)を除いては、ヒトデマン、スターミー共に完全な五放射相称の形をしており、その中心軸は口側-反口側軸と一致する。従って全体の外形としては、ヒトデマンやスターミーのデザインはモチーフのヒトデ(特に腕が5本ある種)の口側-反口側軸を中心軸とした放射相称性をよく反映していると言える。
一方で、ヒトデマン、スターミー共に口側に歩帯溝のような構造や管足らしきものは観察できない。また、反口側の中心付近(ヒトデの盤に対応する部分)に多孔板のような構造は見られない。従って、水管系に関連する形態的特徴の観点からは、ヒトデマンやスターミーのデザインはモチーフのヒトデの特徴を反映しているとは言えない。
ヒドイデとドヒドイデ


ヒドイデとその『進化』先のドヒドイデのモチーフは、タイプ(毒・水)や図鑑説明文の記述からオニヒトデであると推測される(オニヒトデはサンゴを食害し、毒を分泌できる棘を備えている)。
ヒドイデやドヒドイデは頭の上端から触手(ヒドイデ)又は脚(ドヒドイデ)が放射状に(ヒドイデの触手は10本、ドヒドイデの脚は12本)生え、頭の下にさらに体の一部を備える。触手の生え方は頭の中心を通り上下軸に平行な軸を中心軸として放射相称となっているが、この軸は口側-反口側軸とは一致しない。従って、放射相称ではあっても口側-反口側軸を中心軸とする放射相称ではないため、その点ではヒドイデやドヒドイデのデザインはモチーフの特徴を反映しきれていないと言える。
また、ヒドイデ、ドヒドイデ共に触手(脚)の内側(オニヒトデの腕の口側に対応する)や口の周辺には歩帯溝らしき構造は見られず、上側中心部(オニヒトデの盤の反口側に対応する部分)に多孔板らしき構造は見られない。水管系に関連する形態的特徴が見られないという点で、ヒドイデやドヒドイデのデザインはモチーフの特徴を反映しているとは言えない。
ナマコとナマコモチーフのポケモン
ナマコについて
ナマコ綱は棘皮動物の中で唯一進行方向の軸(前後軸)と口側-反口側軸が一致するグループで、一見すると五放射相称であるようには見えないものも多い。しかし、ナマコの体壁の内側には五放射相称に配置された5本の縦走筋がある他、歩帯も(ある場合は)5本走っている。このように外部形態としては観察しづらいものの、ナマコも口側-反口側軸を中心軸とした五放射相称性を備えている。ナマコの口は管足が変化した触手(これを用いて摂食を行う)に囲まれており、その数も5の倍数であることが多い。また、種によっては一部の管足が運動機能を持たない疣足と呼ばれる突起へと変化している。例えば楯手目(潮間帯や浅海帯にいる多くの種はここに含まれる)では、歩帯が接地する側に3本、接地しない側に2本という風に偏っており、接地しない側の2本の歩帯の管足が疣足に変化し、移動には接地する側の3本の歩帯にある管足を用いる(疣足が消失している場合もある)。ナマコの水管系は総排出腔の壁を通じて外部の海水と接続しており、多孔板は持たない(多孔板と相同な穿孔体と呼ばれる構造はあるが、多くのナマコでは外部と繋がっていない)。以上から、触手や歩帯は口側-反口側軸を中心軸とした五放射相称性が(辛うじて)現れる外部形態であると言える。また、水管系に関連する外部形態としては管足やそれが変化した疣足が挙げられる。
補足:ナマコは強いストレスを受けると口又は肛門から消化管、あるいは肛門からキュビエ器官を吐き出す。キュビエ器官は一部の楯手目が持つ器官で、呼吸樹と呼ばれる呼吸器官から繋がる。キュビエ器官は体外に放出されるとナマコ自身以外のものに強く粘着する。
ナマコブシ


ナマコブシのモチーフは、『分類』や名前、図鑑説明文の記述などからナマコであると推測される。
ナマコブシの全体の外形は、スターミーのように明確に口側-反口側軸を中心軸とした五放射相称であるようには見えない。まずは口の形に注目する。ナマコブシの口は「大」の形をしており、中心から放射状に5本の白い線が伸びるデザインとなっている。先述の通りナマコの口は多くの場合5の倍数本の触手によって囲まれており、外部から口側-反口側軸を中心軸とした五放射相称の様子を観察できる部分である。ナマコブシの口には触手のような描写こそないものの、口側-反口側軸を中心軸とした五放射相称性が「大」によって表現されているという点で、ナマコの特徴を反映したデザインであると言える。
次に、ナマコブシの身体の上から出ているピンク色の突起に注目する。この突起はナマコの疣足に対応していると考えられる。先述の通り、浅い海に生息する種を多く含む楯手目の種類では接地しない側の2本の歩帯の管足が疣足に変化しており、疣足は管足と共にナマコの水管系に関連した外部形態である。ピンク色の突起が上側の前後方向に2列存在するというナマコブシのデザインは、モチーフであるナマコの水管系に関連した形態的特徴を、その配置まで含めて非常によく反映していると言える。
ウニとウニモチーフのポケモン
ウニについて
ウニ綱の種は、形態から不正形類とそれ以外(便宜的に正形類と呼ばれる)に大別される。
正形類のウニは上下軸に平行な口側-反口側軸を中心軸とした五放射相称の半球状の殻を持つ。管足の対列とその間の領域をまとめて歩帯と呼び、歩帯は口から放射状に出て殻の表面を上下方向に走る。移動の際の定まった前後は存在しないと言われている。多孔板は肛門とともに殻の頂上付近に存在し、ここを通して外部と水管系の間での海水のやりとりをする。管足は棘と共に移動に用いられる。殻を見るとわかりやすいが、棘の配置も口側-反口側軸を中心軸とした五放射相称になっている。以上から、棘や管足の配置は正形類のウニの口側-反口側軸を中心軸とした五放射相称性がよく現れる外部形態であると言える。また、外部から確認できる水管系の一部として管足およびそれが並ぶ歩帯と、多孔板が挙げられる。
不正形類のウニは移動の際の前後方向が定まっており、左右相称の殻を持つ。しかし内部形態としては環状水管から伸びる5本の放射水管やそれに平行して走る神経系などが観察でき、口側-反口側軸を中心軸とした五放射相称をベースとしていることは正形類と変わらない。口は身体の下側やや前方に、多孔板は頂上部に位置する。口側-反口側軸はやはり上下軸と平行である。歩帯は口から前方に3本、後方に2本伸び、それぞれ頂上へと接続する。頂上付近の歩帯は花のような模様(花紋)を形成する場合がある。管足はタコノマクラ目やカシパン目では歩帯に限らず間歩帯からも出ており、餌を口まで運ぶ際に利用する(身体の移動の際は棘を利用する)。ブンブク目では管足が特殊化し穴を掘ったり餌を集めたりするのに用いられる。以上から、不正形類の口側-反口側軸を中心軸とした五放射相称性を示す外部形態としては口から5本ある程度の放射性をもって伸びる歩帯が挙げられる。また、外部から確認できる水管系の一部としては、管足と多孔板が挙げられる。
バチンウニ



バチンウニは口が前にあり、口側-反口側軸と一致するのは上下軸ではなく前後軸である。口側-反口側軸を中心軸として棘は五放射相称に配置されているようには見えず(一応前から見ると棘は5列あるので歪んだ五放射形に見えなくもないが)、口から伸びる歩帯のような構造も確認できない。上方向や横方向から観察しても棘は五放射相称に配置されていない。従ってヒドイデやドヒドイデのように「中心軸は違うがモチーフの放射相称性が一応反映されている」というわけでもない。バチンウニのデザインは、口側-反口側軸を中心軸とした五放射相称性が全く見られないという点で、モチーフであるウニの特徴を反映しているとは言えない。
バチンウニには歩帯や管足のようなものも確認できない。バチンウニの2対の足(のようなもの)は、ウニの身体に対する管足の太さよりずっと太く、ウニの身体に対する管足の長さよりずっと短いため、足が管足を反映したデザインであるとは断言できない。また、バチンウニのどこを見ても多孔板のような構造は観察できない。従って、バチンウニのデザインは水管系に関連する形態的特徴が見られないという点でモチーフの特徴を反映したものになっているとは言えない。
後書き
以上常体。
棘皮動物「らしさ」を考えた時に「口側-反口側軸を中心軸とする(五)放射相称性があるかどうか」及び「水管系に関する形態的特徴があるかどうか」の2点が重要になると考え考察を行いました。人によってはこれら以外のことに「らしさ」を見出す場合もあると思います。また、棘皮動物についての知識やポケモンの知識で考慮すべき点が考慮できていない、又は間違っている可能性もあります。これらの事情からこの考察も完全なものではありませんが、この記事を見てポケモンのことを考え、それを書いてみようと思う方が増えたら幸いです。
参考文献
各種ポケモンのwiki
https://wiki.ポケモン.com/wiki/リリーラ
https://wiki.ポケモン.com/wiki/ユレイドル
https://wiki.ポケモン.com/wiki/ヒトデマン
https://wiki.ポケモン.com/wiki/スターミー
https://wiki.ポケモン.com/wiki/ヒドイデ
https://wiki.ポケモン.com/wiki/ドヒドイデ
https://wiki.ポケモン.com/wiki/ナマコブシ
https://wiki.ポケモン.com/wiki/バチンウニ
棘皮動物に関するwikipedia
https://ja.wikipedia.org/wiki/ウミユリ綱
https://ja.wikipedia.org/wiki/ウミシダ
https://ja.wikipedia.org/wiki/ウニ
https://ja.wikipedia.org/wiki/棘皮動物
https://ja.wikipedia.org/wiki/水管系
その他のWebページ
https://www.tokyo-zoo.net/topic/topics_detail?kind=news&inst=kasai&link_num=192
https://www.kitasato-u.ac.jp/mb/serc/download/annual_report_2018_02_ver2.pdf
https://ja.wikipedia.org/wiki/刺胞動物
https://ja.wikipedia.org/wiki/有櫛動物
なお、Webページの最終閲覧日はいずれも2021/2/5。
論文
Reich A, Dunn C, Akasaka K, Wessel G (2015) Phylogenomic Analyses of Echinodermata Support the Sister Groups of Asterozoa and Echinozoa. PLoS ONE 10(3): e0119627. doi:10.1371/ journal.pone.0119627
書籍
内田亨 監修(1974). 『動物系統分類学第8巻(中) 棘皮動物』. 中山書店.
本川達雄ら(2003). 『ナマコガイドブック』. 阪急コミュニケーションズ.
佐波征機ら(2002). 『ヒトデガイドブック』. 阪急コミュニケーションズ.
田中颯ら(2019). 『ウニハンドブック』. 文一総合出版.
高橋明義・奥村誠一(2012). 『ナマコ学 -生物・産業・文化-』. 成山堂書店.
池内昌彦ら 監訳(2013). 『キャンベル生物学 原書9版』. 丸善出版.
以上。
技を用いたポケモンの系統分類 Phylogenetic Classification of Pokemon by Using Their Moves
ご挨拶(Greetings)
こんにちは。線虫亭線虫(以下、「著者」)と申します。ブログを書くのは初めてですので至らぬ点が多いかとは思いますが、ご容赦願います。
この記事では、ポケモンをマシン技(技マシンにより覚えることができる技)を用いて系統分類する試みとその結果、考察が書かれています。
以下常体。
- ご挨拶(Greetings)
- 導入(Introduction)
- 手法(Method)
- 結果及び考察(Results and Discussion)
- 補足資料(Supplementary Figures and Table)
- 謝辞(Acknowledgement)
- 参考文献(Reference)
- 追記(Postscript)
導入(Introduction)
「ポケットモンスター」(以下、縮めて「ポケモン」)は今や世界的に有名なゲーム作品のシリーズであり、同時にゲーム内に登場する一群のキャラクターの総称である。ポケモンに対する向き合い方やコンテンツとしての楽しみ方は様々だが、その一つとして、「考察する」ことが挙げられる。今回はポケモンを生物として捉え、その進化について、系統分類学の手法を用いて考察する。
まず、「進化」という言葉について整理し、ポケモンには2種類の「進化」が起きうることを述べる。ポケモンにおいて一般に言及される『進化』とは「同じ個体が、レベルや特定の条件で姿形を変えること」(Nishimu,2015)である。『進化』をすることによって同じ個体でも種族が変わる。これは現在の生物学で用いられる進化の定義とは異なるため、ここでは「同じ個体が、レベルや特定の条件で姿形を変えること」を『進化』とし、現在の生物学において用いられる方の進化(ダーウィンが「変化を伴う継承」と表現したもの)と区別する。ポケモンに進化が起こるためには親から子へと伝わる因子がポケモンに備わっている必要があるが、ポケモンが遺伝子をもつことを示唆する記述は図鑑や公式サイトにもあり、従ってポケモンは進化の必要条件を一つ満たしていると考えられる。また、個体毎に決定される個体値の存在やビビヨンの翅の模様の地域変異、一部のポケモンに見られる性的二型は個体間や地域間、性別間についての種内変異の産物であり、進化において重要な要素である変異がポケモンにも起こることを示している。これらのことを考えれば、ポケモンにおいて進化は起きうるものであり、ポケモンの進化を考えることは有意義であると言える。同種のポケモンが別々の地方の環境に適応した結果とされ、容姿のみならずタイプや特性、種族値、習得技、『進化』先さえも地方間で異なる「リージョンフォーム」の存在やそれに関する記述は、生息環境がポケモンの変化に大きな影響を与えることを強く示唆しており、自然選択によるポケモンの種分化(つまり、ある種の進化)の過程が現に進行していることの興味深い例であると考えられる。
ポケモンの進化を考察する方法の一つとして、ポケモンを「分類する」ことが挙げられる。ポケモンの進化を考察するために分類をするのであれば、その分類が進化の流れ(あるいは進化の系統)を反映したものであることが望ましい。従って、次にどのような分類方法がポケモンの進化を考察する上でより良いかを考える。最も良い方法はポケモンの遺伝物質を直接調べることだろうが、ポケモンの遺伝物質について公式に明確な言及はないため、形質による分類をする他ないと考えられる。形質を用いてポケモンを分類するやり方は大きく分けて以下の3つがあるだろう:A. 図鑑の『分類』の項目による分類、B. 外部形態による分類、C. タマゴグループや特性、タイプ、種族値といったポケモンのそれぞれの種族に固有の性質を利用する分類。これらの3つの分類の仕方について先行研究への言及を交えて述べる。
A. ポケモン図鑑の『分類』の項目による分類
ポケモンが公式に「分類」という言葉を用いている場面の一つがこの項目だが、この項目によるポケモンの分類は著者が調べる限り試みられていない。ポケモンの『分類』はそれぞれのポケモンの外部形態や性質などの要素から一つを取り上げて命名する形式をとっているが、要素の取り上げ方の妥当性には疑問が残る。例えば、ヘラクロスの『分類』は「1ぽんヅノポケモン」であって「かぶとむしポケモン」ではないがカイロスやクワガノンの『分類』は「くわがたポケモン」であって「2ほんヅノポケモン(あるいは、2ほんアゴポケモン)」ではない、同じバリヤードであってもガラルの姿のものは「ダンスポケモン」に『分類』される一方でそれ以外のバリヤードは「バリアーポケモン」に『分類』される、というように取り上げる要素についての統一性が無い場合がしばしばある。取り上げる要素についての統一性がある程度保証されていなければ、『分類』は『分類』をする者の主観に大きく依存してしまうこととなる。従って、『分類』による分類は、『分類』そのものが分類の上で客観的な指標として機能しない可能性があるため難しい。
B. 外部形態による分類
ポケモンの分類方法として外部形態を用いる分類はこれまでしばしば行われてきた。Albertonykus(2017)は「サン・ムーン」までに登場したポケモンについて、それらを実在する生物の系統関係及び公式設定に基づいて「Rock」「Dragon」「Legendary birds」などに分類し系統樹を構築した。「ソード・シールド」までに登場したポケモンについてはたかさおじさん(2020)がポケモンのモチーフに基づく分類を行っており、実在する生物のグループに加えて「ニューエイジ」「擬人化」などといったグループを設けて分類している。外部形態による(系統)分類の際にポケモンのモチーフに対応する実在する生物の分類体系を基にすることは、分類に際して明確な基準を与えられるため有用であると考えられる。しかし、とりわけポケモン間の系統関係を推定する場合においては、実在する生物の系統関係はポケモンの系統関係を反映しているとは限らない(Nishimu,2015)。また、実在する生物の分類体系を基にしない場合、分類に際してどの形質を重視すれば良いかの基準が分類する者の主観に依存することとなり、客観性を十分に保証できないと考えられる。
C. タマゴグループなど種族に固有の性質を利用する分類
ポケモンはゲーム内で種族毎に様々な設定が為されている。公式に明確な言及が無いものも一部存在するものの、分類する際に用いる形質の候補となりうる。Yasuda(2019)は近隣結合法を用いて909種類のポケモンを種族値について階層クラスタリングした。ミケロ(2004)はポケモンをタマゴグループの組み合わせを用いて分類し、それぞれのグループについて実在の生物との関係に関する仮説を立てた。タイプはポケモンが公式に「分類」という言葉を用いて説明している項目の一つだが、これによる分類は著者が調べる限り試みられていない。Nishimu(2015)はタイプ、特性、タマゴグループによる分類はそれぞれ組み合わせが小さすぎると指摘している。また、タイプや特性については特に、ポケモンが技を使用したり(例:「ミラータイプ」、「もりののろい」、「なやみのタネ」は自身や相手のタイプあるいは特性を変える技である)、特性の効果を受けたり(例:「へんげんじざい」、「てんきや」、「ミイラ」は自身や相手のタイプあるいは特性を変える特性である)することによる変化があるため、分類する際の項目として使用するのは適切ではないと考えられる。ポケモンが覚える技による分類はNishimu(2015)によって行われ、「赤・緑」に登場するポケモンが覚える技に対し最大節約法を用いることで系統樹が描かれた。
以上に述べた分類の仕方の中から、ポケモンの進化の系統をできる限り反映した系統樹を作成できるものを採択する。上述の理由からA.及びB.によって分類を行うことは適切ではないだろう。C.において検討されている要素の中で、特にポケモンが覚える技についての分類は効果的であると考えられる。その理由は以下の通り:1.技の性質(タイプ、威力、命中率など)がその技を使う種族に依らず一意に定まるので、多種多様なポケモン全てに共通する形質として扱うことができる、2.1.に述べた共通性がありながら、特定の技を覚えるかどうかについて種族間で異なるため、種族間の多様性の指標としても用いることができる、3.タマゴグループの組み合わせのパターンやタイプの組み合わせのパターンに比べ、覚える技の組み合わせパターンは遥かに多いため詳細な分類を可能にする、4.技は遺伝する形質であると同時に個体間の差をもたらす形質でもある。ここで4.について補足する。ポケモンはタマゴから孵化する際にその親が覚えている技のいくつか(一般に「遺伝技」と呼ばれる)を覚えているが、遺伝技を覚えていない親が持ってきたタマゴから孵化するポケモンは遺伝技を覚えていない(成長しても勝手に習得することもない)。このため、同じ種族でも親が遺伝技を覚えているかどうかで個体によって覚えている技に差が生じるのである(これに対し、種族値などの他の形質は種族に固有である一方で個体間の差はない)。このように、技は系統分類を行う上で扱いやすく、ポケモンの進化を考察する上で重要な特徴を持つ形質であると言える。特に、技マシンで覚えることができる技(以下、「マシン技」)は、技マシンを使用する段階においてポケモンがその技を覚えるかどうかの明確な記述がゲーム内でなされることから、あるポケモンが確実に覚える(あるいは覚えない)技であると考えることが可能である。このため、マシン技はデータの正確さが問われる系統分類に適した性質を持つと言える。
ポケモンの進化の系統をできる限り反映した系統樹を作成するに当たって、形質による闇雲な分類は収斂進化を反映できないという問題を考える必要がある。ポケモンの技もその例外ではないだろう。この問題をできる限り解決するために、作成された系統樹について技以外の収斂が起きにくい形質を頼りに評価する必要がある。この際用いる形質として、タマゴグループが適当であると考えられる。ポケモンがタマゴによって生殖する際に互いのタマゴグループが一致している必要があるという事実は、ポケモンの生殖においてタマゴグループが非常に重要であることを示唆している。種分化の過程が生殖的隔離によって進行するものであることを考えると、生殖可能性に直結する形質が異種間で収斂することは生殖的隔離(あるいは種分化そのもの)に対する逆行であり、考えにくい。従って、タマゴグループは分化することこそあれ収斂することはないという仮定の下で、マシン技によって構築された系統樹をタマゴグループによって評価することにはある程度の合理性があると考えられる。
ポケモンの進化を考察する上で、共通祖先についても考える必要がある。ミュウは公式に「ポケモンの先祖と考える学者がたくさんいる」(ポケモンだいすきクラブ)と明言されている。また、アルセウスはシンオウ神話では宇宙の創造神であると語られている。しかし、これらのポケモンについては研究途上であり、結論に至っていないこともまた公式に名言されている。従って、これらのポケモンを共通祖先と断定して系統樹を構築するのは早計であると言える。系統樹を構築する時、共通祖先の候補として次のような特徴を持つポケモンが挙げられる:A.全てのマシン技を習得できるポケモン、B.どのマシン技も習得できないポケモン。A.の場合、構築された系統樹の背景にある進化の流れは主に「不要な技を喪失しながら環境に適応していく」流れであると考えられる。一方B.の場合、系統樹の背景にある進化の流れは主に「有用な技を獲得しながら環境に適応していく」流れであると考えられる。実在する生物の系統樹の背景にある進化の流れにはB.の方がより近いが、ポケモンの進化の流れが実在の生物のそれと同様であるという仮定をしない限りA.とB.のどちらがより適切かを決めることはできないと考えられる。従って、共通祖先を決定した系統樹の作成は現状困難であるため、無根系統樹を構築するのが望ましいと言える(逆に言えば、ポケモンの進化の流れについて何らかの仮定をすれば有根系統樹を描くことは可能である)。
以上の事情を踏まえ、今回はマシン技を用いたポケモンの系統分類を行い、無根系統樹、及び仮の有根系統樹を描くことを試みた。
手法(Method)

図1:各種ポケモンが覚えるマシン技についてのバイナリーデータを記した表。この表全体を入手できるリンクは補足資料の項目に掲載した。
A. 対象とするポケモン
「赤・緑」から「ウルトラサン・ウルトラムーン」までに登場するポケモンのうち、ウルトラビーストに含まれる種族及び『進化』の余地がある種族(一般に『進化前』と呼ばれる)を除いた426種族を対象とした。ただし、同じ種族であってもリージョンフォームが見られるものはそれぞれの姿を別々にカウントし、ミノマダムについてはそれぞれのミノを別々にカウントしたため、データとしては合計438種類となった。なお、ウルトラビーストは公式にポケモンとして扱われているものの、異世界のポケモンであることが判明しており、ポケモンの進化を考える上でデータに含めることは適切ではないと判断したため、データから除外した。
B. 対象とする技
今回の分類に用いる技は「ウルトラサン・ウルトラムーン」において技マシンに登録されている100種類のマシン技とした。
C. 系統樹作成
まず、RYO(2018)による「第7世代 最終進化一覧」のGoogleスプレッドシートより分類対象となる426種族(438種類)の名前を取り出した。なお、アローラの姿については「[ポケモン名]R」とし、ミノマダムのミノについては「ミノマダム(草)」「ミノマダム(地)」「ミノマダム(鋼)」と表記した。次に、それぞれの種類が覚えるマシン技を「ポケモン徹底攻略」を用いて調べ、覚えない技に0を、覚える技に1を振って「Microsoft Excel(ver.16.35)」で表を作成した(図1)。なお、ロトムはヒートロトムの時にのみ「オーバーヒート」を、トゲキッスは進化前であるトゲチックの時にのみ「スマートホーン」を覚えることができるが、どちらも1を振った。この表を基に、それぞれのポケモンの種類について、技マシン番号の小さい順に0と1を並べた数字の列を作成した。さらにその数字の列の0をG、1をAに置換した文字列を作成した(これらの作業にはExcelの関数を用いた)。この文字列を、解析ソフトで塩基配列を読み込む際に用いられるFASTA形式に書き直し、テキストファイルとして保存した。続いて保存したテキストファイルを分子系統解析ソフト「MEGA7」で読み込み、最大節約法を用いて無根系統樹を作成した。
D. 系統樹分析
作成された無根系統樹に見られるいくつかの単系統群に番号を振って科(Family)とした。なお、Fmailyはそこに含まれるポケモンが40種類を超えないように設定した(一部例外あり)。それぞれのFamilyに対して、以下[]内に定義されるタマゴグループについての「占有率」を求めた。
[N種類のポケモンによって構成されるグループAについて、タマゴグループEをもつ種類の数がNe種類ある時、AにおけるEの占有率=Ne/N]
さらに、マシン技を全く覚えない架空のポケモンを共通祖先と仮定した有根系統樹を上述の無根系統樹から作成し、マシン技ごとに形質の変化が起こった位置を調べ、そのうちFamilyの分化に影響を与えたと考えられるものを有根系統樹内に記した。
結果及び考察(Results and Discussion)



簡略化した無根系統樹を図2、仮の有根系統樹を図3、Family毎の占有率を表1に示した。作成した無根系統樹はFamilyごとに分割し補足図1-17とした。また、分割する前の無根系統樹を入手できるリンクを補足資料の項目に加えた。この無根系統樹に見られる25のFamilyについて、以下にそれぞれの特徴を述べる。
Family1:全5種類。構成するポケモンが覚える技が極端に少ない。タマゴグループも互いに異なっている。
Family2:全7種類。ウルガモスを除いて四足歩行の炎タイプのものからなる。タマゴグループも陸上が7割を占める。ブースターはこのFamilyに含まれる。
Family3:キュウコン及びキュウコンRのみからなる。
Family4:全37種類。草タイプのものが多数を占めるが、一部フェアリータイプが混じる。ナッシーとナッシーRはどちらもこのFamilyに入る。タマゴグループも植物が7割を占める。リーフィアはこのFmailyに含まれる。
Family5:フォレトスとバンバドロのみからなる。タマゴグループは互いに全く異なる。
Family6:全13種類。毒タイプや地面タイプのポケモンを多く含む。特定のタマゴグループが大きく占有しているわけではない。
Family7:カバルドンのみからなる。
Family8:ダグトリオ及びダグトリオRのみからなる。
Family9:ジガルデのみからなる。
Family10:カイロスとヘラクロスのみからなる。
Family11:全26種類。炎タイプとドラゴンタイプのポケモンが多い。ガラガラとガラガラRはどちらもこのFamilyに入る。特定のタマゴグループが大きく占有しているわけではない。
Family12:全11種類。地面タイプと草タイプのポケモンが多い。サンドパンとサンドパンRはどちらもこのFamilyに入る。また、タマゴグループは陸上が5割以上を占める。
Family13:全33種類。伝説のポケモンを除くと全て水タイプまたは氷タイプからなる。タマゴグループは水中1が5割以上を占める。
Family14:全25種類。格闘タイプのものがほとんどで、タマゴグループは人型が5割を占める。
Family15:全31種類。様々なタイプのものから構成されている。ベトベトンとベトベトンRはどちらもこのFamilyに入る。タマゴグループは怪獣及び陸上が4割近くを占めるが、5割を超えるものは無い。
Family16:全33種類。岩タイプや鋼タイプのものが多数含まれる。ゴローニャとゴローニャRはどちらもこのFamilyに入る。タマゴグループは鉱物が4割以上を占める。
Family17:全75種類。水タイプや虫タイプが多く含まれている。構成する種類の数は全てのFamilyの中で最も多い。大雑把に言うと水タイプに対して虫タイプが側系統群を形成する形をとっており、Family17を2つの単系統群に分けるのが難しかったためこのようになっている。タマゴグループは虫と水中1が3割近くを占めるが、4割を超えるものは無い。シャワーズとグレイシアはこのFamilyに含まれる。
Family18:全36種類。飛行タイプのポケモンとエスパータイプのポケモンが多い。タマゴグループは飛行が5割以上を占める。エーフィとニンフィアはこのFamilyに含まれる。
Family19:全27種類。ゴーストタイプや悪タイプのポケモンを多く含む。ペルシアンとペルシアンR、ラッタRはこのFamilyに入る。占有率が4割を超えるタマゴグループは存在しない。ブラッキーはこのFamilyに入る。
Family20:エテボースのみからなる。
Family21:ラッタのみからなる。
Family22:全17種類。電気タイプのポケモンを多く含む。占有率が4割を超えるタマゴグループは存在しない。サンダースはこのFamilyに入る。
Family23:全21種類。電気タイプ(所謂『ピカチュウ枠』)やノーマルタイプのポケモンを多く含む。ライチュウとライチュウRはどちらもこのFamilyに入る。タマゴグループは陸上が6割、妖精が5割を占める。
Family24:ミルホッグのみからなる。
Family25:全28種類。エスパータイプのポケモンを多く含む。占有率が4割を超えるタマゴグループは存在しない。
ここでまず、図3の仮の有根系統樹から推察されるポケモンの仮の進化の道筋を考える。仮の有根系統樹ではメタモンのような「マシン技を一切覚えないポケモン」を共通祖先として設定したが、この理由は主に「形質の獲得をベースに進化を考える方が考察しやすい」ということであり、ミュウ祖先説を否定するものではないことを断っておく。ナマコブシの祖先とFamily2-25の共通祖先とが分岐する前に獲得されていたと考えられる技の中には攻撃技が一切含まれていない。このことは、非常に原始的なポケモンは直接他個体と争うようなことはせず、毒や混乱を用いて間接的に他個体の行動を制限する形で競争をしていたことを示唆している。しかしFamily2-25の共通祖先とナマコブシの祖先が分岐すると、Family2-25の共通祖先側で多数の攻撃技が獲得される。また、「おんがえし」という他個体との協力を示唆する技もこの段階で獲得していることは非常に興味深い。ポケモンの個体間での競争と協力を同時に強めるような選択圧が進化の初期に同時にかかっていたことが伺える。天候操作系の技である「にほんばれ」「あまごい」「すなあらし」もやはり進化の初期に獲得されているが、これらはポケモンが適応しなければならない環境そのものを変化させる技であることを考えると、ある種の環境改変能力の獲得がポケモンの多様化と繁栄をもたらした可能性がある。
続いて、得られた結果からこの系統分類手法の妥当性を検討する。妥当性の検討に際して、まずは確実に系統関係がわかっているポケモン同士の系統樹における位置付けを見る。ポケモンの中には、同じ『進化』前から分岐『進化』して別の種類として扱われるもの(ヤドラン/ヤドキング、所謂『ブイズ』など。以下、「分岐『進化』勢」)や、同一種族ではあるが性別により種類が分けられているもの(ニドキング/ニドクイン、イルミーゼ/バルビート)、リージョンフォームがあるものが存在し、これらに該当するポケモンの組は系統的に互いに最も近いと考えられる。分岐『進化』勢のうち、『進化』先の種類が互いに最も近い関係にあるのは11組(テッカニン/ヌケニンを含む)中3組であった。ただしブイズについては、シャワーズ/グレイシアとエーフィ/ニンフィアはそれぞれ互いに最も近い関係にあった。また、性別により種類が分けられるポケモンはどちらも互いに最も近い関係にあった。リージョンフォームについては、10組中互いに最も近い関係にあるものは5組であった。ただし、これらのうちFamilyを超えて互いに離れている組は『ブイズ』とラッタ/ラッタRのみであった。このような結果は、この方法による系統分類は、既に系統関係が非常に近いとわかっている複数のポケモンについて、それらのポケモンがおおよそ同じFamilyに含まれる程度の精度を有するが、Family未満の関係についての精度までは保障しないことを示唆している。次に、タマゴグループの観点から妥当性を検討する。3種類以上を含む全てのFamilyについて、全ての種類が同一のタマゴグループに属するFamilyは存在しない。従って、今回得られた系統樹は多少なりとも収斂進化が考慮されていないと考えられる。特にFamily1,13,17,26,19,22,25は占有率が4割を超えるタマゴグループが存在せず、Family5は構成される2種類のポケモンの間に共通のタマゴグループがないため、これらのFamilyについて分類の再検討を行う必要がある。ただし、Familyの中にはFamily2や4,23のように一つのタマゴグループが多くを占めるものがあり、それらについてはある程度進化の系統に現れるグループと一致すると考えても構わないと言えるだろう。ここで注意したいのは、タマゴグループによる妥当性の検討も完全なものではないということである。ポケモンの分類学を行う上でタマゴグループがどのように位置付けられ得るかについての考察は、著者の知る限りミケロ(2004)のもの以外存在せず、それもまた実在の生物との関係性を前提とした考察であるから、今回のように実在の生物とは独立にポケモンの進化を考える上では有用ではないだろう。従って、系統樹の妥当性のタマゴグループを用いた考察に先立って、タマゴグループについての新たな整理と考察が必要であると考えられる。
今後の展望として、次の3つが上げられる。1つ目は、先に述べたタマゴグループの整理と考察である。タマゴグループはポケモンの生殖可能性に密接に関連する形質であるが、その種類の少なさからそれのみで分類に用いるには不向きであった。他方、今回のような技による系統分類は妥当性を検証する材料に乏しいという問題がある。技による系統樹と合わせてタマゴグループを考察することは、両者の欠点を補いあい、ポケモンの進化について有益な視点を与えることだろう。2つ目は、新作が登場するたびに生じる技の変化がどれほどの影響をこの分類手法に与えるのかを検討することである。新技の登場によりマシン技の種類が変化することは新作が発売されるごとに起きている。また、「ソード・シールド」の発売に際しては様々な事情からマシン技の種類が変わったり、一部の技が抹消されたりということが起きた。このような変化は今後も避けられないと考えられるが、これに対して系統樹がどのように変化するのかは注目に値する。系統樹の変化の仕方によっては、現時点では判明していない「系統樹を構築する際に適した技のセット(あるいは、系統をより強く反映していると考えられる技のセット)」を解明する手がかりが得られるだろう。
補足資料(Supplementary Figures and Table)
各種ポケモンが覚えるマシン技についてのデータ表は次のリンクからダウンロードできる。
https://docs.google.com/spreadsheets/d/1rtmbCsy3xj4GIz5cKaWt3mB7IwYWpEiwAOQK1Ab-_Ps/edit?usp=sharing
また、作成した無根系統樹の全体の図は次のリンクからダウンロードできる。
https://drive.google.com/file/d/1_YaG2_R1Hyu1IX9OTi8KDyIv8NZxANq8/view?usp=sharing
それぞれのFamily内の系統関係を補足図1-17に示す。

















謝辞(Acknowledgement)
この記事を書くにあたって様々なアドバイスを下さった方々、特に「マシン技を用いたポケモンの系統分類」という概念を思いつき、それを著者に教えて下さった方に感謝申し上げる。
参考文献(Reference)
池内昌彦ら監訳 『キャンベル生物学』第9版 丸善出版
https://wiki.ポケモン.com/wiki/ぶんるい一覧:『分類』について
https://www.slideshare.net/takahironishimu/ss-59418690:Nishimu(2015)による系統分類に関するスライド
http://www2u.biglobe.ne.jp/~endo-c/pokemon/pokelogy/cafetalk/class1.htm:ミケロ(2004)による分類
https://twitter.com/ikiro_pkmn/status/1238386121937018880?s=20:たかさおじさん(2020)による分類
https://www.deviantart.com/albertonykus/art/Pokemon-Phylogeny-212913205:Albertonykus(2017)による系統分類
https://www.pokemon.co.jp/whats/summary/:ポケモン公式によるタイプについての言及
https://vigne-cla.com/16-1/:Yasuda(2019)による階層クラスタリング
https://wiki.ポケモン.com/wiki/進化:『進化』について
https://www.pokemon.jp/special/moyadoga/mew/:ポケモン公式によるミュウについての言及
https://www.pokemon.jp/special/moyadoga/guide/pokemon.html:ポケモン公式によるミュウとアルセウスについての言及
https://lkmmmigapkmn.hatenablog.com/entry/2018/04/03/230958:@Ikm_mm(2018)によるポケモンと遺伝子に関する考察
https://www.pokemon.jp/special/forme/zukan02/:ポケモン公式によるリージョンフォームについての言及
https://tokitamaroku.blog.fc2.com/blog-entry-246.html:今回系統解析の対象としたポケモンのデータ元
https://www.pokemon.co.jp/ex/usum/story/171005_01.html:ポケモン公式によるウルトラビーストについての言及
https://yakkun.com/sm/:タマゴグループやポケモンの画像のデータ元
http://www.nibb.ac.jp/~tomoaki/protocols/genetree/sequence-aquisition.html:FASTA形式について
https://www.megasoftware.net:今回使用した系統解析ソフト「MEGA7」の詳細
https://wiki.ポケモン.com/wiki/タマゴグループ:タマゴグループについて
なお、いずれのWebページも最終閲覧日は2020/6/4。
追記(Postscript)
・見出しの体裁を整え、目次を追加しました。(2021/2/6)
・「補足図(Supplementary Figures)」を「補足資料(Supplementary Figures and Tables)」とし、各種ポケモンが覚えるマシン技に関する表データと、全体の無根系統樹のリンクを追加しました。(2021/2/6)
・「作成された無根系統樹はFamilymごとに分割し補足図1-17とした。」を「作成した無根系統樹はFamilyごとに分割し補足図1-17とした。また、分割する前の無根系統樹を入手できるリンクを補足資料の項目に加えた。」と書き直しました。(2021/2/6)
・図1の説明文に「この表全体を入手できるリンクは補足資料の項目に掲載した。」の一文を加えました。(2021/2/6)
以上。